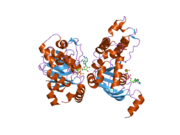Регулятор трансмембранной проводимости при муковисцидозе - Cystic fibrosis transmembrane conductance regulator






Регулятор трансмембранной проводимости при муковисцидозе (CFTR) это мембранный белок и хлоридный канал у позвоночных, что кодируется CFTR ген.[5][6]
Ген CFTR кодирует ABC транспортер -учебный класс ионный канал белок, который проводит хлористый[7] ионы через эпителиальный клеточные мембраны. Мутации гена CFTR, влияющие на функцию канала хлорид-иона, приводят к нарушению регуляции транспорта эпителиальной жидкости в легких, поджелудочной железе и других органах, что приводит к кистозный фиброз. Осложнения включают сгущение слизи в легких при частых респираторных инфекциях и недостаточность поджелудочной железы, приводящую к недоеданию и диабету. Эти состояния приводят к хронической инвалидности и сокращению продолжительности жизни. У пациентов мужского пола прогрессирующая обструкция и разрушение развивающихся семявыносящий проток (семенной канатик) и придаток яичка появляются в результате аномальной внутрипросветной секреции,[8] вызывая врожденное отсутствие семявыносящего протока и мужское бесплодие.
Ген

Ген, кодирующий белок CFTR человека, находится на хромосома 7, на длинном плече в положении q31.2.[6] из базовая пара 116 907 253 для пары оснований 117 095 955. CFTR ортологи[9] происходят в челюстные позвоночные.[10]
В CFTR ген был использован у животных как ядерная ДНК филогенетический маркер.[9] Большие геномные последовательности этого гена были использованы для изучения филогения основных групп млекопитающие,[11] и подтвердил группировку плацентарный заказы в четыре основные клады: Ксенартра, Афротерия, Лавразиатерия, и Euarchonta плюс Glires.
Мутации
Почти 1000 случаев муковисцидоза мутации были описаны.[12] Наиболее распространенная мутация DeltaF508 (ΔF508) возникает в результате делеции (Δ) трех нуклеотидов, что приводит к потере аминокислоты. фенилаланин (F) в 508-м положении белка.[13] В результате белок не складывать нормально и быстрее деградирует. Подавляющее большинство мутаций случаются нечасто. Распространение и частота мутаций варьируются в разных популяциях, что имеет значение для генетического скрининга и консультирования.
Мутации состоят из замен, дупликаций, делеций или укорочений в гене CFTR. Это может привести к тому, что белки могут не функционировать, работать менее эффективно, быстрее разлагаться или присутствовать в недостаточном количестве.[14]
Было высказано предположение, что мутации в гене CFTR могут давать селективное преимущество гетерозиготным индивидуумам. Клетки, экспрессирующие мутантную форму белка CFTR, устойчивы к инвазии со стороны Сальмонелла тифа бактерия, возбудитель брюшной тиф, а мыши, несущие единственную копию мутантного CFTR, устойчивы к диарее, вызванной токсином холеры.[15]
Наиболее частые мутации среди кавказцы находятся:[16]
- ΔF508
- G542X
- G551D
- N1303K
- W1282X
DeltaF508
DeltaF508 (ΔF508), полное имя CFTRΔF508 или же F508del-CFTR (rs113993960 ), представляет собой специфическую мутацию в гене CFTR, включающую удаление из трех нуклеотиды охватывает положения 507 и 508 гена CFTR на хромосоме 7, что в конечном итоге приводит к потере одного кодон для аминокислота фенилаланин (F). Человек с мутацией CFTRΔF508 будет производить аномальный белок CFTR, в котором отсутствует этот остаток фенилаланина и который не может складывать правильно. Этот белок не ускользает от эндоплазматическая сеть для дальнейшей обработки. Наличие двух копий этой мутации (по одной унаследованной от каждого родителя) на сегодняшний день является наиболее частой причиной кистозный фиброз (CF), ответственная за почти две трети случаев во всем мире.[17]
Последствия
Эта секция нужно больше медицинские справки за проверка или слишком сильно полагается на основные источники. (Март 2019 г.) |
Белок CFTR в значительной степени экспрессируется в клетках поджелудочной железы, кишечного и респираторного эпителия и всех экзокринных желез. Когда он правильно свернут, он перемещается к клеточной мембране, где становится трансмембранным белком, ответственным за открытие каналов, которые высвобождают хлористый ионы вне клеток; он также одновременно ингибирует поглощение натрий ионы другим канальным белком. Обе эти функции помогают поддерживать ионный градиент, который вызывает осмос для откачки воды из клеток.[18] Мутация ΔF508 приводит к неправильной укладке CFTR и, в конечном итоге, к его повреждению. деградация в ER. У организмов с двумя дополнениями мутации белок полностью отсутствует на клеточной мембране, и эти важные функции транспорта ионов не выполняются.[19]
Иметь гомозиготный пара генов с мутацией ΔF508 не позволяет белку CFTR занять свое нормальное положение в клеточной мембране. Это вызывает повышенное удержание воды в клетках, соответствующее обезвоживание внеклеточного пространства и связанный с этим каскад воздействия на различные части тела. Эти эффекты включают: более толстый слизистые оболочки в эпителии пораженных органов; закупорка узких дыхательных путей в результате утолщения слизистой и подавление свободного движения мукоцилий; врожденное отсутствие семявыносящего протока из-за увеличения толщины слизи при внутриутробном развитии плода; недостаточность поджелудочной железы из-за закупорки протока поджелудочной железы слизью; и повышенный риск респираторных инфекций из-за накопления густой, богатой питательными веществами слизи, в которой процветают бактерии. Это симптомы кистозный фиброз, генетическое заболевание; однако ΔF508 - не единственная мутация, вызывающая это нарушение.
Быть гетерозиготный перевозчик (имея единственную копию ΔF508) приводит к снижению потерь воды во время понос потому что неисправные или отсутствующие белки CFTR не могут поддерживать стабильные ионные градиенты через клеточные мембраны. Обычно происходит накопление как Cl− и Na+ ионы внутри пораженных клеток, создавая гипотонический раствор вне клеток и заставляет воду диффундировать в клетки за счет осмоса. Несколько исследований показывают, что гетерозиготные носители подвержены повышенному риску развития различных симптомов. Например, было показано, что гетерозиготность по муковисцидозу связана с повышенной реактивностью дыхательных путей, и гетерозиготы могут подвергаться риску плохой легочной функции. Было показано, что гетерозиготы с хрипом подвергаются более высокому риску ухудшения легочной функции или развития и прогрессирования хронических заболеваний. обструктивная болезнь легких. Одного гена муковисцидоза достаточно, чтобы вызвать легкие аномалии легких даже при отсутствии инфекции.[20]
Механизм
Ген CFTR расположен на длинном плече хромосомы 7 в положении q31.2 и в конечном итоге кодирует последовательность из 1480 аминокислот. Обычно три ДНК пар оснований A-T-C (в паре с T-A-G на противоположной цепи) в 507-м положении гена образуют матрицу для кодона мРНК A-U-C для изолейцин, в то время как три пары оснований ДНК T-T-T (спаренные с A-A-A) в соседнем 508-м положении образуют матрицу для кодона U-U-U для фенилаланин.[21] Мутация ΔF508 представляет собой делецию пары C-G из положения 507 вместе с первыми двумя парами T-A из положения 508, оставляя последовательность ДНК A-T-T (спаренную с T-A-A) в положении 507, которое является транскрибируется в кодон мРНК A-U-U. Поскольку A-U-U также кодирует изолейцин, аминокислота в положении 507 не изменяется, и чистый эффект мутации эквивалентен делеции («Δ») последовательности, приводящей к образованию кодона для фенилаланина в положении 508.[22]
Распространенность
ΔF508 присутствует по крайней мере в одной копии хромосомы 7 примерно в одной из 30 Кавказцы. Наличие мутации на обеих копиях вызывает аутосомно-рецессивный болезнь муковисцидоз. Ученые подсчитали, что первоначальная мутация произошла более 52000 лет назад в Северной Европа. Молодой аллель возраст может быть следствием прошлого выбора. Одна из гипотез о том, почему вредная мутация поддерживается естественным отбором, заключается в том, что одна копия может оказывать положительный эффект, уменьшая потерю воды во время холера, хотя введение патогенных Холерный вибрион в Европу не попал до конца 18 века.[23] Другая теория утверждает, что носители CF (гетерозиготы по ΔF508) более устойчивы к брюшной тиф, поскольку было показано, что CFTR действует как рецептор для Сальмонелла тифа бактерии попадают в эпителиальные клетки кишечника.[24]
Муковисцидоз Гетерозиготы ΔF508 могут быть перепредставлены среди людей с астма и может иметь более низкую функцию легких, чем у лиц, не являющихся носителями.[25][26] Носители одной мутации МВ имеют более высокую распространенность хронических заболеваний. риносинусит чем население в целом.[27] Примерно 50% случаев муковисцидоза в Европа происходят из-за гомозиготных мутаций ΔF508 (это сильно варьируется в зависимости от региона),[28] в то время как частота аллеля ΔF508 составляет около 70%.[29] Остальные случаи вызваны более чем 1500 другими мутациями, включая R117H, 1717-1G> A и 2789 + 56G> A. Эти мутации в сочетании друг с другом или даже с одной копией ΔF508 могут вызывать симптомы МВ. Генотип не сильно коррелирует с тяжестью CF, хотя определенные симптомы связаны с определенными мутациями.
Структура

Ген CFTR составляет примерно 189 kb в длину, с 27 экзоны и 26 интроны.[30] CFTR - это гликопротеин с 1480 г. аминокислоты. Белок состоит из пяти доменов. Есть два трансмембранных домена, каждый с шестью участками альфа спирали. Каждый из них подключен к нуклеотид-связывающий домен (NBD) в цитоплазме. Первый NBD связан со вторым трансмембранным доменом регуляторным «R» -доменом, который является уникальной особенностью CFTR и не присутствует в других Автовозы ABC. Ионный канал открывается только тогда, когда его R-домен фосфорилирован PKA и АТФ привязан к NBD.[31] В карбоксильный терминал белка прикреплен к цитоскелет по ПДЗ -взаимодействующий домен.[32] Показанная структура (PDB # 1XMI) показывает гомопентамерную сборку мутировавшего NBD1, первого нуклеотид-связывающего домена (NBD1) переносчика.
Расположение и функции

CFTR функционирует как фосфорилирование и АТФ -закрытый анион канал, увеличивая проводимость для некоторых анионы (например, Cl−) стечь их электрохимический градиент. АТФ-управляемый конформационные изменения в CFTR открывать и закрывать ворота, чтобы позволить трансмембранному потоку анионов вниз по их электрохимический градиент.[33] Это в отличие от других ABC белки, в котором управляемые АТФ конформационные изменения подпитывают транспорт субстрата через клеточные мембраны. По сути, CFTR - это ионный канал, который превратился в «сломанный» ABC транспортер что протекает, когда открыт конформация.
CFTR имеют два трансмембранных домена, каждый из которых связан с нуклеотид-связывающим доменом. CFTR также содержит другой домен, называемый регуляторным доменом. Другие члены надсемейства переносчиков ABC участвуют в поглощении питательных веществ прокариотами или в экспорте различных субстратов у эукариот. ABC-транспортеры эволюционировали, чтобы преобразовывать свободную энергию гидролиза АТФ в восходящее движение субстратов через клеточную мембрану. У них есть две основные конформации: одна, где сайт связывания груза обращена к цитозолю или обращена внутрь (без АТФ), а другая - где она обращена наружу (связана с АТФ). АТФ связывается с каждым нуклеотид-связывающим доменом, что приводит к последующей димеризации NBD, что приводит к перестройке трансмембранных спиралей. Это изменяет доступность места привязки груза из положения, обращенного внутрь, на положение, обращенное наружу. Связывание АТФ и последующий гидролиз приводят к альтернативному открытию места связывания груза, обеспечивая однонаправленную транспортировку груза против электрохимический градиент. В CFTR чередование конформации, обращенной внутрь, к конформации, обращенной наружу, приводит к стробированию канала. В частности, димеризация NBD (чему способствует связывание АТФ) сопряжена с переходом в обращенную наружу конформацию, в которой образуется открытый трансмембранный путь для анионов. Последующий гидролиз (в каноническом активном сайте, сайте 2, включая мотивы Уокера NBD2) дестабилизирует димер NBD и способствует возвращению к обращенной внутрь конформации, в которой путь проникновения анионов закрыт.[33]
CFTR обнаружен в эпителиальных клетках многих органов, включая легкое, печень, поджелудочная железа, пищеварительный тракт, а женский[34] и мужчина репродуктивный трактаты.[35][36]
В дыхательных путях легких CFTR наиболее сильно экспрессируется редкими специализированными клетками, называемыми легочные ионоциты.[37][38][39] В коже CFTR сильно выражен в сальный и эккрин потовые железы.[40] В эккриновых железах CFTR располагается на апикальной мембране эпителиальных клеток, составляющих проток этих потовых желез.[40]
Обычно белок позволяет перемещать хлористый и тиоцианат[41] ионы (с отрицательным зарядом) из эпителиальной клетки в поверхностную жидкость дыхательных путей и слизь. Положительно заряженные ионы натрия следуют пассивно, увеличивая общий электролит концентрация в слизи, в результате чего вода выходит из клетки через осмос.
В эпителиальных клетках с подвижными ресничками, выстилающими бронх и яйцевод, CFTR располагается на апикальной клеточной мембране, но не на ресничках.[34] В отличие, ENaC (Эпителиальный натриевый канал) располагается по всей длине ресничек.[34]
В потовые железы, дефектный CFTR приводит к снижению транспорта хлорида натрия и натрия тиоцианат[42] в реабсорбирующем протоке и, следовательно, более соленый пот. Это основа клинически важного потовый тест за кистозный фиброз часто используется в диагностике при генетическом скрининге.[43]
Взаимодействия
Было показано, что регулятор трансмембранной проводимости при муковисцидозе взаимодействовать с:
Подавляется препаратом против диареи. крофелемер.
Связанные условия
- Врожденное двустороннее отсутствие семявыносящего протока: Мужчины с врожденным двусторонним отсутствием семявыносящий проток чаще всего имеют легкую мутация (изменение, которое допускает частичную функцию гена) в одной копии гена CFTR и мутацию, вызывающую муковисцидоз, в другой копии CFTR.
- Кистозный фиброз: Обнаружено более 1800 мутаций в гене CFTR.[57] но большинство из них не были связаны с муковисцидозом.[58] Большинство этих мутаций либо заменяют одну аминокислота (строительный блок белков) для другой аминокислоты в белке CFTR или удалить небольшое количество ДНК в гене CFTR. Наиболее распространенная мутация, называемая ΔF508, представляет собой делецию (Δ) одной аминокислоты (фенилаланин) в положении 508 в белке CFTR. Этот измененный белок никогда не достигает клеточной мембраны, потому что он разрушается вскоре после образования. Все вызывающие заболевание мутации в гене CFTR препятствуют правильному функционированию канала, что приводит к блокированию движения соли и воды в клетки и из них. В результате этой блокировки клетки, выстилающие проходы в легких, поджелудочной железе и других органах, производят аномально густую липкую слизь. Эта слизь закупоривает дыхательные пути и железы, вызывая характерные признаки и симптомы муковисцидоза. Кроме того, только жидкую слизь можно удалить с помощью реснички; густая слизь не может, поэтому она задерживает бактерии, вызывающие хронические инфекции.
- Холера: АДФ-рибозилирование вызванный холерный токсин приводит к увеличению производства циклический AMP что, в свою очередь, открывает канал CFTR, что приводит к избыточной секреции Cl−. Na+ и H2O следовать за Cl− в тонкий кишечник, что приводит к обезвоживанию и потере электролитов.[59]
Мишень для лекарства
CFTR был мишень для наркотиков в попытках найти способы лечения связанных состояний. Ивакафтор (торговое наименование Калидеко, разработанная как VX-770) это препарат, средство, медикамент одобрен FDA в 2012 году для людей с кистозный фиброз у которых есть специфические мутации CFTR.[60][61] Ивакафтор был разработан Vertex Pharmaceuticals в сочетании с Фонд кистозного фиброза и является первым лекарством, которое лечит основную причину, а не симптомы заболевания.[62] Названный «самым важным новым лекарством 2012 года»,[63] и "чудо-наркотик"[64] это один из самых дорогих препаратов, стоимость которого превышает АМЕРИКАНСКИЙ ДОЛЛАР$ 300000 в год, что привело к критике Vertex за высокую стоимость.
Рекомендации
- ^ а б c ГРЧ38: Ансамбль выпуск 89: ENSG00000001626 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000041301 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Гэдсби, округ Колумбия, Вергани П., Чанади Л. (март 2006 г.). «Белок ABC превратил канал хлорида, отказ которого вызывает муковисцидоз». Природа. 440 (7083): 477–83. Bibcode:2006Натура.440..477Г. Дои:10.1038 / природа04712. ЧВК 2720541. PMID 16554808.
- ^ а б Коллинз Ф, Ромменс Дж. М., Ианнуцци М. С., Керем Б., Драмм М. Л., Мелмер Дж., Дин М., Розмахель Р., Коул Дж. Л., Кеннеди Д., Хидака Н. (сентябрь 1989 г.). «Идентификация гена муковисцидоза: ходьба и прыжки по хромосомам». Наука. 245 (4922): 1059–65. Bibcode:1989Научный ... 245.1059R. Дои:10.1126 / science.2772657. PMID 2772657.
- ^ Риордан Дж. Р., Ромменс Дж. М., Керем Б., Алон Н., Розмахель Р., Гржелчак З., Зеленски Дж., Лок С., Плавшич Н., Чоу Дж. Л. (сентябрь 1989 г.). «Идентификация гена муковисцидоза: клонирование и характеристика комплементарной ДНК». Наука. 245 (4922): 1066–73. Bibcode:1989Научный ... 245.1066R. Дои:10.1126 / science.2475911. PMID 2475911.
- ^ Marcorelles P, Gillet D, Friocourt G, Ledé F, Samaison L, Huguen G, Ferec C (март 2012 г.). «Экспрессия белка регулятора трансмембранной проводимости при муковисцидозе в системе мужских выводных протоков во время развития». Патология человека. 43 (3): 390–7. Дои:10.1016 / j.humpath.2011.04.031. PMID 21840567.
- ^ а б «Филогенетический маркер OrthoMaM: кодирующая последовательность CFTR». Архивировано из оригинал на 2016-03-02. Получено 2010-03-12.
- ^ Дэвис, Р. Конрой, С.Дж .; Дэвис, WL; Поттер, IC; Rrezise, Ann EO (19–23 июня 2005 г.). «Эволюция и регуляция гена кистозного фиброза» (доклад конференции). Конференция по молекулярной биологии и эволюции (MBE05). Получено 28 июля 2014.
- ^ Prasad AB, Allard MW, Green ED (сентябрь 2008 г.). «Подтверждение филогении млекопитающих с использованием больших наборов сравнительных данных последовательностей». Молекулярная биология и эволюция. 25 (9): 1795–808. Дои:10.1093 / молбев / msn104. ЧВК 2515873. PMID 18453548.
- ^ «Клиническая и функциональная интерпретация CFTR (CFTR2): история списка вариантов CFTR2». Фонд CF США, Университет Джона Хопкинса, Центр муковисцидоза при Детской больнице в Торонто. Получено 2 августа 2017.[постоянная мертвая ссылка ]
- ^ Гуимбеллот, Дженнифер; Шарма, Джьоти; Роу, Стивен М. (ноябрь 2017 г.). «На пути к инклюзивной терапии с модуляторами CFTR: прогресс и проблемы». Детская пульмонология. 52 (S48): S4 – S14. Дои:10.1002 / ppul.23773. ISSN 1099-0496. ЧВК 6208153. PMID 28881097.
- ^ Роу С.М., Миллер С., Соршер Э.Дж. (май 2005 г.). "Кистозный фиброз". Медицинский журнал Новой Англии. 352 (19): 1992–2001. Дои:10.1056 / NEJMra043184. PMID 15888700.
- ^ Кавич С.М., Фрем Э.Дж., Сегал А.С. (1999). «Тематические исследования холеры: уроки истории медицины и естествознания». Йельский журнал биологии и медицины. 72 (6): 393–408. ЧВК 2579035. PMID 11138935.
- ^ Араужо Ф.Г., Новаес ФК, Сантос Н.П., Мартинс В.К., Соуза С.М., Сантос С.Е., Рибейру-дос-Сантос АК (январь 2005 г.). «Распространенность мутаций deltaF508, G551D, G542X и R553X среди пациентов с муковисцидозом на севере Бразилии». Бразильский журнал медицинских и биологических исследований = Revista Brasileira de Pesquisas Medicas e Biologicas. 38 (1): 11–5. Дои:10.1590 / S0100-879X2005000100003. PMID 15665983.
- ^ Бобадилла, JL; Мацек-младший, М; Хорошо, JP; Фаррелл, PM (2002). «Муковисцидоз: всемирный анализ мутаций CFTR - корреляция с данными о заболеваемости и применение для скрининга». Человеческая мутация. 19 (6): 575–606. Дои:10.1002 / humu.10041. PMID 12007216.
- ^ Веркман, А.С .; Песня, Y .; Тиагараджа, Дж. Р. (2003). «Роль жидкости на поверхности дыхательных путей и подслизистых желез при муковисцидозе легких». Американский журнал физиологии. Клеточная физиология. 284 (1): C2 – C15. Дои:10.1152 / ajpcell.00417.2002. PMID 12475759.
- ^ «Направления исследования кистозного фиброза». Национальный институт диабета, болезней органов пищеварения и почек (NIDDK).
- ^ Маурья, Нутан; Авасти, Шалли; Дикшит, Пратибха (апрель 2012 г.). «Ассоциация мутации гена CFTR с бронхиальной астмой» (PDF). Индийский J Med: 469–478. Получено 4 февраля, 2015.
- ^ Отчет CCDS для согласованного CDS: Отчет для CCDS5773.1 (текущая версия) NCBI
- ^ Bartoszewski, R.A .; Яблонский, М .; Bartoszewska, S .; Стивенсон, Л .; Dai, Q .; Kappes, J .; Collawn, J.F .; Бебок, З. (13 июля 2010 г.). «Синонимичный однонуклеотидный полиморфизм в ΔF508 CFTR изменяет вторичную структуру мРНК и экспрессию мутантного белка». Журнал биологической химии. 285 (37): 28741–28748. Дои:10.1074 / jbc.M110.154575. ЧВК 2937902. PMID 20628052.
- ^ "Re: Есть ли связь между муковисцидозом и холерой?".
- ^ Pier, G.B .; Затирка, М .; Заиди, Т .; Meluleni, G .; Mueschenborn, S.S .; Banting, G .; Ratcliff, R .; Evans, M.J .; Колледж, W.H. (7 мая 1998 г.). "Сальмонелла тифа использует CFTR для проникновения в эпителиальные клетки кишечника ». Природа. 393 (6680): 79–82. Bibcode:1998Натура.393 ... 79П. Дои:10.1038/30006. PMID 9590693. S2CID 5894247.
- ^ Даль, Мортен; Nordestgaard, Børge G .; Ланге, Питер; Тыбьерг-Хансен, Энн (8 января 2001 г.). «Пятнадцатилетнее наблюдение за функцией легких у лиц, гетерозиготных по делеции фенилаланина-508 при муковисцидозе». Аллергия Клин Иммунол. 107 (5): 818–823. Дои:10.1067 / mai.2001.114117. PMID 11344348.
- ^ Даль, М; Tybjaerg-Hansen, A; Lange, P; Nordestgaard, BG (27 июня 1998 г.). «Гетерозиготность DeltaF508 при муковисцидозе и предрасположенность к астме». Ланцет. 351 (9120): 1911–3. Дои:10.1016 / с0140-6736 (97) 11419-2. PMID 9654257. S2CID 22970136.
- ^ Ван, Синьцзин; Ким, Жан; Маквильямс, Рита; Катание, Гарри Р. (март 2005 г.). «Повышенная распространенность хронического риносинусита у носителей мутации муковисцидоза». Хирургия головы и шеи Arch Otolaryngol. 131 (3): 237–40. Дои:10.1001 / archotol.131.3.237. PMID 15781764.
- ^ Годовой отчет ECFS: что это значит для Великобритании Муковисцидоз Trust
- ^ Morral, N; Bertranpetit, J; Эстивилл, X; Nunes, R; Casals, T; Хименес, Дж; Ангеличева, Д (1994). «Происхождение основной мутации кистозного фиброза (ΔF508) в европейских популяциях». Природа Генетика. 7 (2): 169–75. Дои:10.1038 / ng0694-169. PMID 7920636. S2CID 38005421.
- ^ База данных мутаций кистозного фиброза. «Последовательность геномной ДНК». Архивировано из оригинал на 22.08.2016. Получено 2013-04-06.
- ^ Шеппард Д. Н., валлийский MJ (январь 1999 г.). «Строение и функция хлоридного канала CFTR». Физиологические обзоры. 79 (1 приложение): S23-45. Дои:10.1152 / физрев.1999.79.1.S23. PMID 9922375.
- ^ а б Short DB, Trotter KW, Reczek D, Kreda SM, Bretscher A, Boucher RC, Stutts MJ, Milgram SL (июль 1998 г.). «Апикальный белок PDZ прикрепляет регулятор трансмембранной проводимости при муковисцидозе к цитоскелету». Журнал биологической химии. 273 (31): 19797–801. Дои:10.1074 / jbc.273.31.19797. PMID 9677412.
- ^ а б Csanády, L; Vergani, P; Гэдсби, округ Колумбия (1 января 2019 г.). «Структура, стробирование и регулирование анионного канала CFTR» (PDF). Физиологические обзоры. 99 (1): 707–738. Дои:10.1152 / Physrev.00007.2018. PMID 30516439.
- ^ а б c Энука Ю., Ханукоглу И., Эдельхейт О, Вакнин Х., Ханукоглу А. (март 2012 г.). «Эпителиальные натриевые каналы (ENaC) равномерно распределены на подвижных ресничках в яйцеводе и дыхательных путях». Гистохимия и клеточная биология. 137 (3): 339–53. Дои:10.1007 / s00418-011-0904-1. PMID 22207244. S2CID 15178940.
- ^ Шарма С., Ханукоглу А., Ханукоглу И. (2018). «Локализация эпителиального натриевого канала (ENaC) и CFTR в зародышевом эпителии яичка, клетках Сертоли и сперматозоидах». Журнал молекулярной гистологии. 49 (2): 195–208. Дои:10.1007 / s10735-018-9759-2. PMID 29453757. S2CID 3761720.CS1 maint: несколько имен: список авторов (связь)
- ^ Шарма С., Ханукоглу I (2019). «Картирование участков локализации эпителиального натриевого канала (ENaC) и CFTR в сегментах придатка яичка млекопитающих». Журнал молекулярной гистологии. 50 (2): 141–154. Дои:10.1007 / s10735-019-09813-3. PMID 30659401. S2CID 58026884.
- ^ «Исследование CF обнаруживает новые клетки, называемые ионоцитами, несущие высокие уровни гена CFTR». Новости муковисцидоза сегодня. 3 августа 2018.
- ^ Montoro DT, Haber AL, Biton M, Vinarsky V, Lin B, Birket SE, Yuan F, Chen S, Leung HM, Villoria J, Rogel N, Burgin G, Tsankov AM, Waghray A, Slyper M, Waldman J, Nguyen L , Dionne D, Rozenblatt-Rosen O, Tata PR, Mou H, Shivaraju M, Bihler H, Mense M, Tearney GJ, Rowe SM, Engelhardt JF, Regev A, Rajagopal J (август 2018 г.). «Пересмотренная иерархия эпителия дыхательных путей включает ионоциты, экспрессирующие CFTR». Природа. 560 (7718): 319–324. Bibcode:2018Натура.560..319М. Дои:10.1038 / s41586-018-0393-7. ЧВК 6295155. PMID 30069044.
- ^ Плассхаерт Л.В., Жилионис Р., Чу-Винг Р., Савова В., Кнехр Дж., Рома Г., Кляйн А.М., Яффе А.Б. (август 2018 г.). «Одноклеточный атлас эпителия дыхательных путей показывает богатые CFTR легочные ионоциты». Природа. 560 (7718): 377–381. Bibcode:2018Натура.560..377П. Дои:10.1038 / s41586-018-0394-6. ЧВК 6108322. PMID 30069046.
- ^ а б Ханукоглу И., Боггула В.Р., Вакнин Х., Шарма С., Клейман Т., Ханукоглу А. (июнь 2017 г.). «Экспрессия эпителиального натриевого канала (ENaC) и CFTR в эпидермисе человека и эпидермальных придатках». Гистохимия и клеточная биология. 147 (6): 733–748. Дои:10.1007 / s00418-016-1535-3. PMID 28130590. S2CID 8504408.
- ^ Moskwa P, Lorentzen D, Excoffon KJ, Zabner J, McCray PB, Nauseef WM, Dupuy C, Bánfi B (январь 2007 г.). «Новая система защиты дыхательных путей хозяина дефектна при муковисцидозе». Американский журнал респираторной медицины и реанимации. 175 (2): 174–83. Дои:10.1164 / rccm.200607-1029OC. ЧВК 2720149. PMID 17082494.
- ^ Xu Y, Szép S, Lu Z (декабрь 2009 г.). «Антиоксидантная роль тиоцианата в патогенезе муковисцидоза и других заболеваний, связанных с воспалением». Труды Национальной академии наук Соединенных Штатов Америки. 106 (48): 20515–9. Bibcode:2009PNAS..10620515X. Дои:10.1073 / pnas.0911412106. ЧВК 2777967. PMID 19918082.
- ^ Ёнэй Ю., Танака М., Одзава И., Миядзаки К., Цукада Н., Инада С., Инагаки Ю., Миямото К., Судзуки О., Окава Х. (апрель 1992 г.). «Первичная гепатоцеллюлярная карцинома с тяжелой гипогликемией: участие инсулиноподобных факторов роста». Печень. 12 (2): 90–3. Дои:10.1111 / j.1600-0676.1992.tb00563.x. PMID 1320177.
- ^ Чжан Х., Питерс К.В., Сунь Ф., Марино С.Р., Ланг Дж., Бургойн Р.Д., Фриззелл Р.А. (август 2002 г.). «Белок цепочки цистеина взаимодействует с регулятором трансмембранной проводимости при муковисцидозе и модулирует его созревание». Журнал биологической химии. 277 (32): 28948–58. Дои:10.1074 / jbc.M111706200. PMID 12039948.
- ^ Ченг Дж., Мойер Б.Д., Милевски М., Лоффинг Дж., Икеда М., Микл Дж. Э., Каттинг Г.Р., Ли М., Стэнтон Б.А., Гуджино В.Б. (февраль 2002 г.). «Связанный с Гольджи белок домена PDZ модулирует экспрессию трансмембранного регулятора муковисцидоза на плазматической мембране». Журнал биологической химии. 277 (5): 3520–9. Дои:10.1074 / jbc.M110177200. PMID 11707463.
- ^ а б Gentzsch M, Cui L, Mengos A, Chang XB, Chen JH, Riordan JR (февраль 2003 г.). «PDZ-связывающий хлоридный канал ClC-3B локализуется в Гольджи и ассоциируется с белками PDZ, взаимодействующими с регулятором трансмембранной проводимости при муковисцидозе». Журнал биологической химии. 278 (8): 6440–9. Дои:10.1074 / jbc.M211050200. PMID 12471024.
- ^ Ван С., Юэ Х, Дерин Р. Б., Гуджино В. Б., Ли М. (сентябрь 2000 г.). «Дополнительный белок способствует взаимодействию CFTR-CFTR, молекулярному механизму усиления активности хлоридных каналов». Клетка. 103 (1): 169–79. Дои:10.1016 / S0092-8674 (00) 00096-9. PMID 11051556. S2CID 16697781.
- ^ Liedtke CM, Yun CH, Kyle N, Wang D (июнь 2002 г.). «Протеинкиназа C эпсилон-зависимая регуляция трансмембранного регулятора муковисцидоза включает связывание с рецептором для активированной C-киназы (RACK1) и связывание RACK1 с фактором регуляции обмена Na + / H +». Журнал биологической химии. 277 (25): 22925–33. Дои:10.1074 / jbc.M201917200. PMID 11956211.
- ^ а б Пак М., Ко С.Б., Чой Дж. Й., Муаллем Дж., Томас П. Дж., Пушкин А., Ли М. С., Ким Дж. Ю., Ли М. Г., Муаллем С., Курц И. (декабрь 2002 г.). «Регулятор трансмембранной проводимости при муковисцидозе взаимодействует с изоформой 3 переноса HCO3-спасения человека Na + -HCO3- и регулирует их активность». Журнал биологической химии. 277 (52): 50503–9. Дои:10.1074 / jbc.M201862200. PMID 12403779.
- ^ а б Кормет-Бояка Э., Ди А., Чанг С.И., Нарен А.П., Туссон А., Нельсон Д.Д., Кирк К.Л. (сентябрь 2002 г.). «Каналы хлорида CFTR регулируются комплексом SNAP-23 / синтаксин 1A». Труды Национальной академии наук Соединенных Штатов Америки. 99 (19): 12477–82. Bibcode:2002PNAS ... 9912477C. Дои:10.1073 / пнас.192203899. ЧВК 129470. PMID 12209004.
- ^ Хегедюс Т., Сесслер Т., Скотт Р., Телин В., Бакос Е., Варади А., Сабо К., Хомоля Л., Милграм С.Л., Саркади Б. (март 2003 г.). «С-концевое фосфорилирование MRP2 модулирует его взаимодействие с белками PDZ». Сообщения о биохимических и биофизических исследованиях. 302 (3): 454–61. Дои:10.1016 / S0006-291X (03) 00196-7. PMID 12615054.
- ^ Ван С., Рааб Р.В., Шац П.Дж., Гуджино В.Б., Ли М. (май 1998 г.). «Консенсус связывания пептидов домена NHE-RF-PDZ1 соответствует С-концевой последовательности регулятора трансмембранной проводимости при муковисцидозе (CFTR)». Письма FEBS. 427 (1): 103–8. Дои:10.1016 / S0014-5793 (98) 00402-5. PMID 9613608. S2CID 20803242.
- ^ Мойер Б.Д., Духайм М., Шоу С., Дентон Дж., Рейнольдс Д., Карлсон К.Х., Пфайффер Дж., Ван С., Микл Дж. Э., Милевски М., Каттинг Г. Р., Гуджино В. Б., Ли М., Стэнтон Б. «PDZ-взаимодействующий домен регулятора трансмембранной проводимости при муковисцидозе необходим для функциональной экспрессии в апикальной плазматической мембране». Журнал биологической химии. 275 (35): 27069–74. Дои:10.1074 / jbc.M004951200. PMID 10852925.
- ^ Hall RA, Ostedgaard LS, Premont RT, Blitzer JT, Rahman N, Welsh MJ, Lefkowitz RJ (июль 1998 г.). «С-концевой мотив, обнаруженный в бета2-адренергическом рецепторе, рецепторе P2Y1 и регуляторе трансмембранной проводимости при муковисцидозе, определяет связывание с семейством факторов регуляции обменника Na + / H + белков PDZ». Труды Национальной академии наук Соединенных Штатов Америки. 95 (15): 8496–501. Bibcode:1998PNAS ... 95.8496H. Дои:10.1073 / пнас.95.15.8496. ЧВК 21104. PMID 9671706.
- ^ Сунь Ф., Хуг М.Дж., Леварчик С.М., Юн С.Х., Брэдбери Н.А., Фриззелл Р.А. (сентябрь 2000 г.). «E3KARP опосредует ассоциацию эзрина и протеинкиназы A с регулятором трансмембранной проводимости при муковисцидозе в клетках дыхательных путей». Журнал биологической химии. 275 (38): 29539–46. Дои:10.1074 / jbc.M004961200. PMID 10893422.
- ^ Нарен А.П., Нельсон Д.Д., Се В., Джовов Б., Певснер Дж., Беннетт М.К., Бенос Д.Д., Квик М.В., Кирк К.Л. (ноябрь 1997 г.). «Регулирование хлоридных каналов CFTR с помощью синтаксина и изоформ Munc18». Природа. 390 (6657): 302–5. Bibcode:1997Натура.390..302Н. Дои:10.1038/36882. PMID 9384384. S2CID 4395005.
- ^ Иган М.Э. (март 2016 г.). «Генетика муковисцидоза: клиническое значение». Клиники грудной медицины. 37 (1): 9–16. Дои:10.1016 / j.ccm.2015.11.002. PMID 26857764.
- ^ Де Бок, Крис; Амарал, Маргарида Д (август 2016 г.). «Прогресс в терапии муковисцидоза». Ланцет респираторной медицины. 4 (8): 662–674. Дои:10.1016 / S2213-2600 (16) 00023-0. PMID 27053340.
- ^ Тиагараджа Дж. Р., Веркман А. С. (сентябрь 2012 г.). «Ингибиторы CFTR для лечения диарейного заболевания». Клиническая фармакология и терапия. 92 (3): 287–90. Дои:10.1038 / clpt.2012.114. ЧВК 3643514. PMID 22850599.
- ^ Джонс А.М., Хелм Дж. М. (октябрь 2009 г.). «Новые методы лечения муковисцидоза». Наркотики. 69 (14): 1903–10. Дои:10.2165/11318500-000000000-00000. PMID 19747007. S2CID 23344660.
- ^ McPhail GL, Clancy JP (апрель 2013 г.). «Ивакафтор: первая терапия, действующая на основную причину муковисцидоза». Наркотики сегодняшнего дня. 49 (4): 253–60. Дои:10.1358 / dot.2013.49.4.1940984. PMID 23616952.
- ^ «Исследование фазы 3 VX-770 показывает заметное улучшение функции легких у людей с муковисцидозом с мутацией G551D». Пресс-релиз. Фонд кистозного фиброза. 2011-02-23.
- ^ Herper M (27 декабря 2012 г.). «Самый важный новый препарат 2012 года». Forbes.
- ^ Nocera J (18 июля 2014 г.). "Лекарство за 300 000 долларов". Нью-Йорк Таймс.
дальнейшее чтение
- Kulczycki LL, Kostuch M, Bellanti JA (январь 2003 г.). «Клиническая перспектива муковисцидоза и новые генетические данные: связь мутаций CFTR с проявлениями генотипа-фенотипа». Американский журнал медицинской генетики. Часть А. 116A (3): 262–7. Дои:10.1002 / ajmg.a.10886. PMID 12503104. S2CID 9245855.
- Ванкеерберген А., Куппенс Х, Кассиман Дж. Дж. (Март 2002 г.). «Трансмембранный регулятор проводимости при муковисцидозе: интересный белок с плейотропными функциями». Журнал муковисцидоза. 1 (1): 13–29. Дои:10.1016 / S1569-1993 (01) 00003-0. PMID 15463806.
- Цуй LC (1992). «Мутации и вариации последовательности, обнаруженные в гене регулятора трансмембранной проводимости муковисцидоза (CFTR): отчет Консорциума генетического анализа муковисцидоза». Человеческая мутация. 1 (3): 197–203. Дои:10.1002 / humu.1380010304. PMID 1284534. S2CID 35904538.
- Макинтош I, Cutting GR (июль 1992 г.). «Муковисцидоз, регулятор трансмембранной проводимости и этиология и патогенез муковисцидоза». Журнал FASEB. 6 (10): 2775–82. Дои:10.1096 / fasebj.6.10.1378801. PMID 1378801. S2CID 24932803.
- Драмм М.Л., Коллинз Ф.С. (1993). «Молекулярная биология муковисцидоза». Молекулярно-генетическая медицина. 3: 33–68. Дои:10.1016 / b978-0-12-462003-2.50006-7. ISBN 9780124620032. PMID 7693108.
- Керем Б., Керем Э. (1996). «Молекулярные основы изменчивости муковисцидоза». Европейский журнал генетики человека. 4 (2): 65–73. Дои:10.1159/000472174. PMID 8744024. S2CID 41476164.
- Девидас С., Гуджино В.Б. (октябрь 1997 г.). «CFTR: области, структура и функции». Журнал биоэнергетики и биомембран. 29 (5): 443–51. Дои:10.1023 / А: 1022430906284. PMID 9511929. S2CID 6000695.
- Нагель Дж. (Декабрь 1999 г.). «Дифференциальная функция двух нуклеотидсвязывающих доменов на регуляторе трансмембранной проводимости при муковисцидозе». Biochimica et Biophysica Acta (BBA) - Биомембраны. 1461 (2): 263–74. Дои:10.1016 / S0005-2736 (99) 00162-5. PMID 10581360.
- Бойл МП (2000). «Уникальные проявления и хронические осложнения муковисцидоза у взрослых: учат ли они нас чему-нибудь о МВТР?». Респираторные исследования. 1 (3): 133–5. Дои:10.1186 / rr23. ЧВК 59552. PMID 11667976.
- Грегер Р., Шрайбер Р., Молл М., Висснер А., Хопф А., Бриль М., Блайх М., Варт Р., Кунцельманн К. (2001). «Муковисцидоз и МВТР». Pflügers Archiv. 443 Дополнение 1: S3-7. Дои:10.1007 / s004240100635. PMID 11845294. S2CID 8057614.
- Брэдбери Н.А. (2001). «Сигнальные каскады cAMP и CFTR: есть ли еще чему поучиться?». Pflügers Archiv. 443 Приложение 1: S85-91. Дои:10.1007 / s004240100651. PMID 11845310. S2CID 19373036.
- Дахан Д., Эвагелидис А., Ханрахан Дж. В., Хинксон Д. А., Цзя Ю., Ло Дж., Чжу Т. (2001). «Регулирование канала CFTR путем фосфорилирования». Pflügers Archiv. 443 Дополнение 1: S92-6. Дои:10.1007 / s004240100652. PMID 11845311. S2CID 8144727.
- Кон Дж. А., Ноун П. Г., Джоуэлл П. С. (сентябрь 2002 г.). «Идиопатический панкреатит, связанный с CFTR: сложное наследование и идентификация гена-модификатора». Журнал следственной медицины. 50 (5): 247S – 255S. Дои:10.1136 / jim-50-suppl5-01. PMID 12227654. S2CID 34017638.
- Шварц М. (февраль 2003 г.). «[Ген муковисцидоза трансмембранного регулятора проводимости (CFTR): мутации и клинические фенотипы]». Ugeskrift для Laeger. 165 (9): 912–6. PMID 12661515.
- Вонг Л.Дж., Альпер О.М., Ван Б.Т., Ли М.Х., Ло С.И. (июль 2003 г.). «Две новые нулевые мутации у тайваньского пациента с муковисцидозом и обзор мутаций CFTR в Восточной Азии». Американский журнал медицинской генетики. Часть А. 120А (2): 296–8. Дои:10.1002 / ajmg.a.20039. PMID 12833420. S2CID 41060230.
- Куппенс Х., Кассиман Дж. Дж. (Октябрь 2004 г.). «Мутации и полиморфизмы CFTR при мужском бесплодии». Международный журнал андрологии. 27 (5): 251–6. Дои:10.1111 / j.1365-2605.2004.00485.x. PMID 15379964.
- Кон Дж. А., Митчелл Р. М., Джоуэлл П. С. (март 2005 г.). «Влияние муковисцидоза и мутаций гена PSTI / SPINK1 на предрасположенность к хроническому панкреатиту». Клиники лабораторной медицины. 25 (1): 79–100. Дои:10.1016 / j.cll.2004.12.007. PMID 15749233.
- Южный К.В., Пекхэм Д. (2004). «Установление диагноза муковисцидоз». Хроническое респираторное заболевание. 1 (4): 205–10. Дои:10.1191 / 1479972304cd044rs. PMID 16281647.
- Кандула Л., Уиткомб, округ Колумбия, Лоу, штат Мэн (июнь 2006 г.). «Генетические проблемы при детском панкреатите». Текущие отчеты гастроэнтерологии. 8 (3): 248–53. Дои:10.1007 / s11894-006-0083-8. PMID 16764792. S2CID 23606613.
- Марсет Б., Бойнаемс Дж. М. (декабрь 2006 г.). «Взаимосвязь между регулятором трансмембранной проводимости муковисцидоза, внеклеточными нуклеотидами и муковисцидозом». Фармакология и терапия. 112 (3): 719–32. Дои:10.1016 / j.pharmthera.2006.05.010. PMID 16828872.
- Wilschanski M, Durie PR (август 2007 г.). «Паттерны заболевания желудочно-кишечного тракта в зрелом возрасте, связанные с мутациями в гене CFTR». Кишечник. 56 (8): 1153–63. Дои:10.1136 / gut.2004.062786. ЧВК 1955522. PMID 17446304.
внешняя ссылка
- Статья в GeneReviews / NCBI / NIH / UW о заболеваниях, связанных с МВТР - муковисцидозе (МВ, муковисцидоз) и врожденном отсутствии семявыносящего протока (CAVD)
- Белок-регулятор трансмембранной проводимости при муковисцидозе
- База данных мутаций генов человека - записи CFTR
- База данных мутаций кистозного фиброза
- Информация CFTR Национальной лаборатории Окриджа
- CFTR в OMIM (Национальный центр биотехнологической информации)
- Обзор всей структурной информации, доступной в PDB за UniProt: P13569 (Регулятор трансмембранной проводимости при муковисцидозе человека) в PDBe-KB.
- Обзор всей структурной информации, доступной в PDB за UniProt: P26361 (Регулятор трансмембранной проводимости при муковисцидозе мышей) на PDBe-KB.
PDB галерея | |
|---|---|
|